
Go to any reptile show or zoo, and I can guarantee you will find herpetoculture’s future generation pointing at a reptile or amphibian on displa
Go to any reptile show or zoo, and I can guarantee you will find herpetoculture’s future generation pointing at a reptile or amphibian on display and asking their parents, “Mommy, Daddy is that snake/frog/ lizard poisonous?” Everyone has to walk before they can run, and everyone working with reptiles today—even venomous reptiles—was likely once that child.
Information on Venomous Snakes
Herping for Venomous Snakes in Uganda

Sean M. Perry
Lateral view of the king cobra (Ophiophagus hannah); the oval-shaped venom gland is behind the eye in elapids such as this.
Each year while I was attending veterinary school, my school held an open house and invited boy scouts, girl scouts and other community organizations from around Southern California. My first year contributing to this event, we created a reptile house, which became the most popular stop for children because they were able to interact with many different reptiles.
One section of the reptile exhibit was dedicated to tri-color colubrids, including California mountain kingsnakes (Lampropeltis zonata) and Arizona mountain kingsnakes (L. pyromelanea), and I would frequently observe the previously described scenario with children asking if the snakes were poisonous. One of our goals was to help educate them by describing the coral snake mimicry system and the difference between venomous and poisonous reptiles, and then allow each child to interact with non-venomous mimics. Over the years, children would return to the reptile house and use the terms “venomous” and “poisonous” correctly and in the right context, and I even observed several teaching other children about the proper usage of the words.
Our goal with this article is to clarify and educate on how to use the words venomous and poisonous in the appropriate context when discussing reptiles and amphibians. In addition, we will discuss the major venom and poison delivery systems that have been described in reptiles and amphibians.
Defining Terms
“Venomous” can be used to describe animals that deliver venom using a specialized structure to impair another organism. Venom is a potentially toxic secretion produced by an organism to the impairment of another organism. For example, in snakes, the specialized structure used for venom delivery in elapids and vipers are front fangs, while non-front-fanged colubrids have deeply grooved maxillary teeth that facilitate venom delivery during a bite.

Sean M. Perry
The areas indicated on these two photos show where the venom glands are located on the most venomous snake in the world, the inland taipan (Oxyuranus microlepidotus).
One principal difference between venom and poison is that venomous animals actively deliver their toxic secretions, which then impair the target species. Poisonous animals rely on passive means of afflicting another organism. Poisonous reptiles produce secretions that are absorbed by a predator’s epithelial lining (e.g., skin or gastrointestinal tract), leading to an alteration in the physiology of the consuming organism. For example, in the wild, poison dart frogs of the family Dendrobatidae excrete toxins from granular glands embedded in their skin, which are used as an anti-predation tool. Most poisonous reptiles deliver their toxin passively, either by being ingested or secreting a toxic substance that is absorbed by a predator.
Venom Delivery Systems in Snakes
Approximately 3,000 extant advanced snakes belong to the superfamily Colubroidea, including vipers and elapids. Natural selection allowed for the evolution of venom systems from specialized salivary gland excretions and modifications of teeth in order to improve the snakes’ abilities to capture and subdue prey. Venom’s principal function is to aid in capturing prey and potentially enhancing digestion. Additionally, it can be used in a defensive manner.
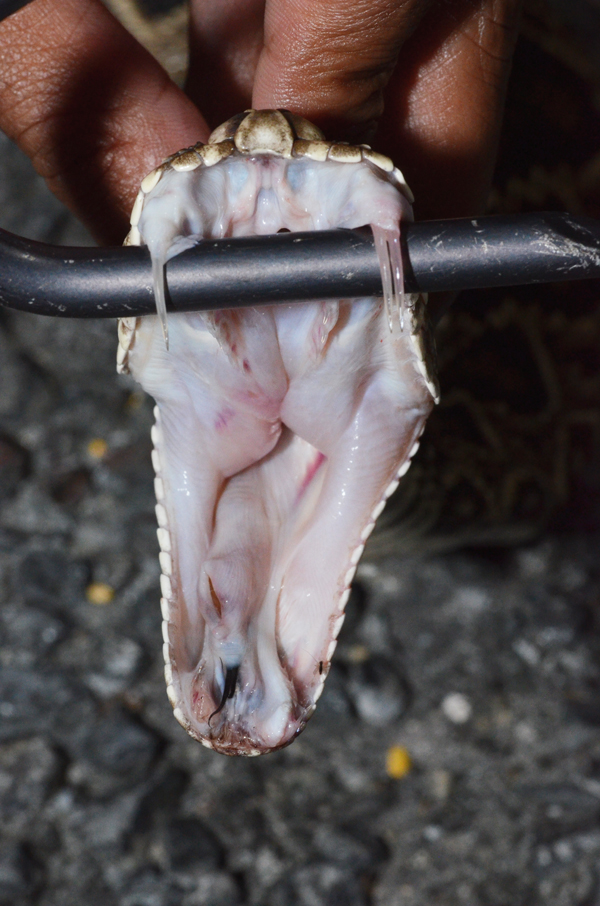
Sean M. Perry
Being displayed are the two large, kinetic fangs of an eastern diamondback rattlesnake (Crotalus adamanteus).
Venom injected into prey reduces the contact time between predator and prey item. This is vital because fractious and biting prey can be injurious or even lethal to a snake. Reduced prey handling time reduces the use of energy involved in subduing the prey and decreases injury to the snake.
Classical venomous species (viperids [solenoglyphous]) and elapids [proteroglyphous]). Classical venomous species are found on every continent except Antarctica. Elapid and viper venom is delivered in a similar fashion, with major anatomical differences in the delivery apparatus. The functional structure of the venom gland and front-fanged dentition differs considerably compared to other venomous reptile species.
These genera possess venom glands, providing a storage system that allows venom to pool within the glandular system. Muscles attach to the gland and exert high-pressure, facilitating rapid delivery of a significant volume of pre-stored venom through the associated venom ducts and fangs. In elapids, two oval-shaped glands are located behind the eyes. The venom is released from the glandular cells upon muscular contraction, coursing through an accessory gland and through the anterior fixed fangs.
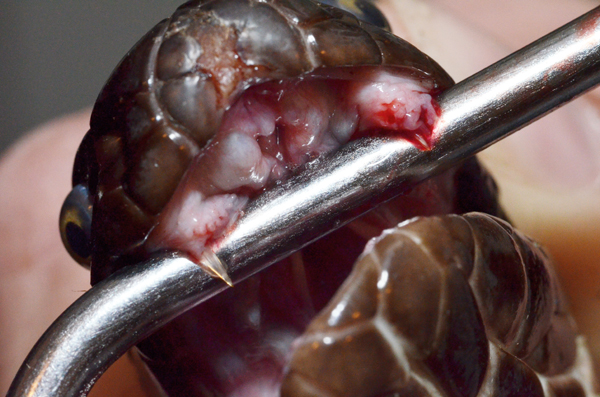
SEAN M. PERRY
Two small fixed fangs are present in the oral cavity of this Pakistani cobra (Naja naja karachiensis).
In comparison, viperids have two large, triangular glands behind the eyes. Upon envenomation, muscles compress the main venom gland and the venom travels through a primary duct into an accessory venom gland. From there, it passes through a secondary duct and into the large, tubular kinetic fangs within the anterior oral cavity, and into the prey.
Atractaspidinae species
Atractaspis species are primarily found in Africa and the Middle East. They are unique in their fang structure and venom gland, possessing elongated venom glands that have been documented to extend much further caudally than the skull compared to vipers and elapids. Some species, such as the stiletto snakes, posses enlarged canaliculated fangs that are freely rotating on the upper jaw. This adaptation allows for these species to envenomate from lateral movements, projecting their fangs out in a lateral fashion rather than in an open mouth delivery as in most venomous reptiles. Manual handling of these snakes is not recommended because of their highly mobile lateral fangs and the high risk of becoming envenomated.
Rear-fanged venomous species (Colubridae [opisthoglyphous])
Venom gland structure in viperids and elapids is significantly different than the jaw and gland structure of colubrids, known as the Duvernoy’s gland. The Duvernoy’s gland shows significant variation in size between species, lacks any significant storage capacity and contains a duct system different from front-fanged snakes. Additionally, there are no direct muscle attachments to pressurize the gland, resulting in a low-pressure secretion-injection system.

Sean M. Perry
The circled area indicates where the Duvernoy’s gland is located in
this false water cobra (Hydrodynastes gigas).
Many rear-fanged snakes have multiple enlarged teeth, either at the mid point or the caudal aspect of the upper jaw, adapted for enhanced conduction of secretions into the inflicted bite wound. The presence of a grooved “rear fang” is not strictly required to introduce Duvernoy’s secretions into a bite wound. Compared to elapids and vipers, the quantity of venom introduced into an envenomation site is considerably less.
Rather than envenomate using the viper/elapid method of muscular attachments and compression of the gland to inject stored venom, colubrids envenomate using a combination of stimulation from the nervous system and pressurization due to anatomical relationships between the venom ducts and surrounding structures. The released secretion from the Duvernoy’s gland is conveyed by a duct into the oral cavity and adjacent teeth. It is then conducted by capillary action along the surface of the teeth, often facilitated by dental grooves, where it is not injected, but rather inoculated, into the bite wound of the prey or predator.
Venomous Lizards
Gila monsters (Heloderma suspectum) and beaded lizards (H. horridum) deliver venom through paired venom glands that empty through ducts at the base of venom-conducting teeth. Venom is produced and stored in venom glands in the lower jaw. Heloderm venom glands are not surrounded by compressive muscles to assist in envenomation; instead, tension within the gland produced by jaw movements propels venom toward the teeth.

SEAN M. PERRY
The venom gland of the Gila monster (Heloderma suspectum) is located along the side of the lower jaw.
A series of small folds and grooved tissue in this area serves as a temporary reservoir for the venom and facilitates the delivery from venom gland to tooth. Capillary action created by the grooved teeth ensures venom delivery into a bite wound. Helodermatids do not specialize in envenomating large quantities of venom during a bite; rather, their venom delivery system is structured to quickly and effectively transfer venom into the bites they deliver.
Non-helodermatid Lizard Species
Including non-helodermatid lizard species within this article is controversial. Some great research has been done, specifically in regard to the Komodo dragon (Varanus komododesis), to prove that some varanid species are truly venomous. In addition, there are reports of toxic components and transcripts encoding several classes of toxin-like proteins in the oral secretions of non-helodermatid lizards, such as iguanids, agamids and other varanid species. However, the jury within the scientific community is still out.

SEAN M. PERRY
The venomous characteristics of the Komodo dragon (Varanus komodoensis) have been studied, but according to the authors, the jury is still out on whether or not the lizard should be considered truly venomous.
Using the definition provided within this article, it is still debatable whether or not to call the Komodo dragon “venomous.” It does possess specialized glands that secrete toxic substances functionally similarly to the Duvernoy’s gland; however, no current evidence exists demonstrating these proteins are introduced into prey at levels significant enough to produce rapid impairment of the prey item in the wild or experimentally.
Poisonous Amphibians
In amphibians, granular skin glands are responsible for producing chemicals that range from irritating to lethal. These glands are evenly spread across the dorsal surface or concentrated in large glandular masses on each individual species.
Poison Dart Frogs (Dendrobatidae)
Poison dart frogs are popular species kept within the reptile trade despite their known toxicity. It has been well documented that poison dart frogs accumulate and sequester some of their skin toxins from dietary sources, and they produce some of the most toxic alkaloid poisons known. Dendrobatid frogs raised on a diet of insects derived from forest leaf litter accumulate toxins in their skin, whereas frogs raised on fruit flies do not. Possible sources of dendrobatid toxins include ants (pyrrolizidines and indolizidines), beetles (coccinellines) and small millipedes (pyrrolizidine oximes).

SEAN M. PERRY
The toxins of poison dart frogs, such as this blue dart frog (Dendrobates tinctorius), are excreted from glands embedded in their skin.
Their toxins are excreted from glands embedded in the skin and function as a co-evolutionary, anti-predator mechanism in conjunction with their bright coloration (aposematic coloration). The toxins secreted onto the skin must be ingested or cross through skin via a cut in order to be effective. Tribes in South America traditionally use the frogs’ skin secretions to aid in hunting and immobilizing prey items. Wild-caught specimens can maintain toxicity for some time, so appropriate care should be taken when handling such animals, especially newly acquired specimens. After a long-term captive diet of fruit flies, the frogs will eventually lose their toxicity.
Bufo Species
Toads of the genus Bufo have evolved specialized parotid glands that secrete serous milky white poisonous material used as an anti-predator defense mechanism. They typically require significant stimulation to produce the secretion and are only employed in a defensive manner in conjunction with defensive behaviors. The milky substance that is released contains one of the most toxic chemicals characterized in nature, tetrodotoxin, which blocks nerve signals within the body and can even kill humans when ingested. Predators that come into contact with tetrodotoxin will feel its effects within minutes.

SEAN M. PERRY
The circled area indicates the location of the parotid gland in this southern toad (Anaxyrus [Bufo] terrestris).
Newts (Taricha and Cynops spp.)
Newts, like Bufo toads, also have parotid and granular glands that form secretions, resulting in a protective covering that makes the newt unpalatable and toxic to any predator that attempts to consume it. Again, tetrodotoxin is employed.
Older newts are typically more toxic than younger specimens; however, the tetrodotoxin isolated from the eggs of some Taricha species is far more potent. Certain populations of Taricha species are also more toxic, and they have co-evolved with predators in an evolutionary arms race. Male newts, too, tend to be more toxic than females.
Snakes That Are Both Venomous and Poisonous
Certain species and populations of garter snake (Thamnophis sirtalis, couchii and atratus) retain their toxins from their prey items, making these snakes poisonous in addition to possessing a functional Duvernoy’s gland. In these species, the venom produced from the Duvernoy’s gland is toxic to amphibians and other small animals, but it is not harmful to humans. Despite their venom not being harmful to humans, these garter snake species retain tetrodotoxin from rough-skinned newt and American toad prey items. Thamnophis species have developed a resistance to ingesting the tetrodotoxin and using it themselves as a defense while decreasing their own anti-predator behaviors.

SEAN M. PERRY
Garter snakes (Thamnophis spp.), though not dangerous to humans, could actually be considered both venomous and poisonous.
The tiger keelback (Rhabdophis tigrinus) possesses two nuchal glands behind the head that accumulate several steroid toxins. This species has both a Duvernoy’s gland and a poison-containing nuchal gland, making it another snake that is both venomous and poisonous. The nuchal gland stores toxins that are consumed along with toad prey items. Therefore, the toxins that are secreted from the nuchal gland are primarily bufandienolides, which are analogous to the batrachotoxins that have been similarly isolated from a few genera of poison dart frogs. These toxins primarily affect the heart, and the function of the nuchal gland is primarily anti-predator in nature.
It’s difficult to cover such a broad topic such as venom/poison delivery systems in an article of this length, especially when there are many variations within a group of animals as diverse as reptiles and amphibians. For additional in-depth information, read books such as Mackessy’s Handbook of Venoms and Toxins of Reptiles, and Fry’s soon-to-be released, Venomous Reptiles and Their Toxins: Evolution, Pathophysiology and Biodiscovery.
In addition, there’s a lot of literature on these subjects within the scientific community, with research being published often on venom systems, venom composition, toxins and novel species. We encourage you to learn as much as possible about these wondrous species, and with any luck, the information presented in this article has piqued the interest of future generations of herpetologists and herpetoculturists in venomous and poisonous reptiles.
Sean Perry is an associate veterinarian at Marathon Veterinary Hospital in the Florida Keys, with a special interest in reptile medicine and surgery. He also works at The Turtle Hospital in Marathon, Fla. Douglas Mader, MS, DVM, DABVP (C/F, R/A), DECZM (HERPETOLOGY), is a graduate of the University of California, Davis. He owns the Marathon Veterinary Hospital in the Conch Republic, and is a world-renowned lecturer, author and editor. He sits on the review boards of several scientific and veterinary journals.
